10.3 Stems
Mary Ann Clark; Jung Choi; and Matthew Douglas
Learning Objectives
By the end of this section, you will be able to do the following:
- Describe the main function and basic structure of stems
- Compare and contrast the roles of dermal tissue, vascular tissue, and ground tissue
- Distinguish between primary growth and secondary growth in stems
- Summarize the origin of annual rings
- List and describe examples of modified stems
Stems are a part of the shoot system of a plant. They may range in length from a few millimeters to hundreds of meters, and also vary in diameter, depending on the plant type. Stems are usually above ground, although the stems of some plants, such as the potato, also grow underground. Stems may be herbaceous (soft) or woody in nature. Their main function is to provide support to the plant, holding leaves, flowers, and buds; in some cases, stems also store food for the plant. A stem may be unbranched, like that of a palm tree, or it may be highly branched, like that of a magnolia tree. The stem of the plant connects the roots to the leaves, helping to transport absorbed water and minerals to different parts of the plant. It also helps to transport the products of photosynthesis, namely sugars, from the leaves to the rest of the plant.
Plant stems, whether above or below ground, are characterized by the presence of nodes and internodes (see figure below). Nodes are points of attachment for leaves, aerial roots, and flowers. The stem region between two nodes is called an internode. The stalk that extends from the stem to the base of the leaf is the petiole. An axillary bud is usually found in the axil—the area between the base of a leaf and the stem—where it can give rise to a branch or a flower. The apex (tip) of the shoot contains the apical meristem within the apical bud.

Stem Anatomy
The stem and other plant organs arise from the ground tissue and are primarily made up of simple tissues formed from three types of cells: parenchyma, collenchyma, and sclerenchyma cells (see figure below).
Parenchyma cells are the most common plant cells. They are found in the stem, the root, the inside of the leaf, and the pulp of the fruit. Parenchyma cells are responsible for metabolic functions, such as photosynthesis, and they help repair and heal wounds. Some parenchyma cells also store starch.

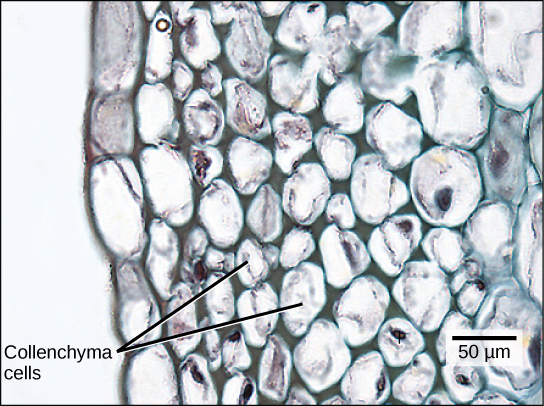
Collenchyma cells are elongated cells with unevenly thickened walls (see figure below). They provide structural support, mainly to the stem and leaves. These cells are alive at maturity and are usually found below the epidermis. The “strings” of a celery stalk are an example of collenchyma cells.
Sclerenchyma cells also provide support to the plant, but unlike collenchyma cells, many of them are dead at maturity. There are two types of sclerenchyma cells: fibers and sclereids. Both types have secondary cell walls that are thickened with deposits of lignin, an organic compound that is a key component of wood. Fibers are long, slender cells; sclereids are smaller-sized. Sclereids give pears their gritty texture. Humans use sclerenchyma fibers to make linen and rope (see figure below).

Like the rest of the plant, the stem has three tissue systems: dermal, vascular, and ground tissue. Each is distinguished by characteristic cell types that perform specific tasks necessary for the plant’s growth and survival.
Dermal Tissue
The dermal tissue of the stem consists primarily of epidermis, a single layer of cells covering and protecting the underlying tissue. Woody plants have a tough, waterproof outer layer of cork cells commonly known as bark, which further protects the plant from damage. Epidermal cells are the most numerous and least differentiated of the cells in the epidermis. The epidermis of a leaf also contains openings known as stomata, through which the exchange of gases takes place. Two cells, known as guard cells, surround each leaf stoma, controlling its opening and closing and thus regulating the uptake of carbon dioxide and the release of oxygen and water vapor (see figure below). Trichomes are hair-like structures on the epidermal surface. They help to reduce transpiration (the loss of water by aboveground plant parts), increase solar reflectance, and store compounds that defend the leaves against predation by herbivores.

Vascular Tissue
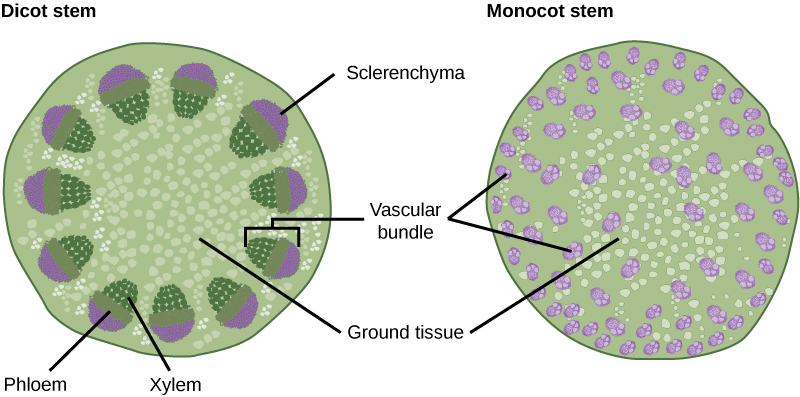
The xylem and phloem that make up the vascular tissue of the stem are arranged in distinct strands called vascular bundles, which run up and down the length of the stem. When the stem is viewed in cross section, the vascular bundles of dicot stems are arranged in a ring. In plants with stems that live for more than one year, the individual bundles grow together and produce the characteristic growth rings. In monocot stems, the vascular bundles are randomly scattered throughout the ground tissue (see figure below).
Xylem tissue has three types of cells: xylem parenchyma, tracheids, and vessel elements. The latter two types conduct water and are dead at maturity. Tracheids are xylem cells with thick secondary cell walls that are lignified. Water moves from one tracheid to another through regions on the side walls known as pits, where secondary walls are absent. Vessel elements are xylem cells with thinner walls; they are shorter than tracheids. Each vessel element is connected to the next by means of a perforation plate at the end walls of the element. Water moves through the perforation plates to travel up the plant.
Phloem tissue is composed of sieve-tube cells, companion cells, phloem parenchyma, and phloem fibers. A series of sieve-tube cells (also called sieve-tube elements) are arranged end to end to make up a long sieve tube, which transports organic substances such as sugars and amino acids. The sugars flow from one sieve-tube cell to the next through perforated sieve plates, which are found at the end junctions between two cells. Although still alive at maturity, the nucleus and other cell components of the sieve-tube cells have disintegrated. Companion cells are found alongside the sieve-tube cells, providing them with metabolic support. The companion cells contain more ribosomes and mitochondria than the sieve-tube cells, which lack some cellular organelles.
Ground Tissue
Ground tissue is mostly made up of parenchyma cells, but may also contain collenchyma and sclerenchyma cells that help support the stem. The ground tissue towards the interior of the vascular tissue in a stem or root is known as pith, while the layer of tissue between the vascular tissue and the epidermis is known as the cortex.
Growth in Stems
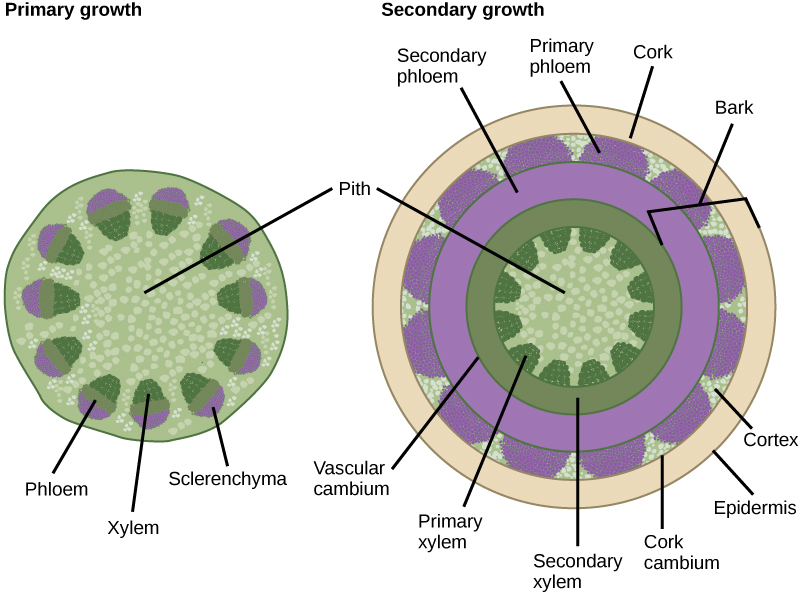
Growth in plants occurs as the stems and roots lengthen. Some plants, especially those that are woody, also increase in thickness during their life span. The increase in length of the shoot and the root is referred to as primary growth and is the result of cell division in the shoot apical meristem. Secondary growth is characterized by an increase in the thickness or girth of the plant and is caused by cell division in the lateral meristem. The figure below shows the areas of primary and secondary growth in a plant. Herbaceous plants mostly undergo primary growth, with hardly any secondary growth or increase in thickness. Secondary growth or “wood” is noticeable in woody plants; it occurs in some dicots but occurs very rarely in monocots.
Some plant parts, such as stems and roots, continue to grow throughout a plant’s life: a phenomenon called indeterminate growth. Other plant parts, such as leaves and flowers, exhibit determinate growth, which ceases when a plant part reaches a particular size.
Primary Growth
Most primary growth occurs at the apices, or tips, of stems and roots. Primary growth is a result of rapidly dividing cells in the apical meristems at the shoot tip and root tip. Subsequent cell elongation also contributes to primary growth. The growth of shoots and roots during primary growth enables plants to continuously seek water (roots) or sunlight (shoots).
The influence of the apical bud on overall plant growth is known as apical dominance, which diminishes the growth of axillary buds that form along the sides of branches and stems. Most coniferous trees exhibit strong apical dominance, thus producing the typical conical Christmas tree shape. If the apical bud is removed, then the axillary buds will start forming lateral branches. Gardeners make use of this fact when they prune plants by cutting off the tops of branches, thus encouraging the axillary buds to grow out, giving the plant a bushy shape.
Link to Learning
Watch this BBC Nature video showing how time-lapse photography captures plant growth at high speed
Secondary Growth
The increase in stem thickness that results from secondary growth is due to the activity of the lateral meristems, which are lacking in herbaceous plants. Lateral meristems include the vascular cambium and, in woody plants, the cork cambium (see figure below). The vascular cambium is located just outside the primary xylem and to the interior of the primary phloem. The cells of the vascular cambium divide and form secondary xylem (tracheids and vessel elements) to the inside, and secondary phloem (sieve elements and companion cells) to the outside. The thickening of the stem that occurs in secondary growth is due to the formation of secondary phloem and secondary xylem by the vascular cambium, plus the action of cork cambium, which forms the tough outermost layer of the stem. The cells of the secondary xylem contain lignin, which provides hardiness and strength.
In woody plants, cork cambium is the outermost lateral meristem. It produces cork cells (bark) containing a waxy substance known as suberin that can repel water. The bark protects the plant against physical damage and helps reduce water loss. The cork cambium also produces a layer of cells known as phelloderm, which grows inward from the cambium. The cork cambium, cork cells, and phelloderm are collectively termed the periderm. The periderm substitutes for the epidermis in mature plants. In some plants, the periderm has many openings, known as lenticels (see figure below), which allow the interior cells to exchange gases with the outside atmosphere. This supplies oxygen to the living and metabolically active cells of the cortex, xylem, and phloem.

Annual Rings
The activity of the vascular cambium gives rise to annual growth rings. During the spring growing season, cells of the secondary xylem have a large internal diameter and their primary cell walls are not extensively thickened. This is known as early wood, or spring wood. During the fall season, the secondary xylem develops thickened cell walls, forming late wood, or autumn wood, which is denser than early wood. This alternation of early and late wood is due largely to a seasonal decrease in the number of vessel elements and a seasonal increase in the number of tracheids. It results in the formation of an annual ring, which can be seen as a circular ring in the cross section of the stem. An examination of the number of annual rings and their nature (such as their size and cell wall thickness) can reveal the age of the tree and the prevailing climatic conditions during each season (see figure below).

Stem Modifications
Some plant species have modified stems that are especially suited to a particular habitat and environment. A rhizome is a modified stem that grows horizontally underground and has nodes and internodes. Vertical shoots may arise from the buds on the rhizome of some plants, such as ginger and ferns. Corms are similar to rhizomes, except they are more rounded and fleshy (such as in gladiolus). Corms contain stored food that enables some plants to survive the winter. Stolons are stems that run almost parallel to the ground, or just below the surface, and can give rise to new plants at the nodes. Runners are a type of stolon that runs above the ground and produces new clone plants at nodes at varying intervals: strawberries are an example. Tubers are modified stems that may store starch, as seen in the potato (Solanum sp.). Tubers arise as swollen ends of stolons and contain many adventitious or unusual buds (familiar to us as the “eyes” on potatoes). A bulb, which functions as an underground storage unit, is a modification of a stem that has the appearance of enlarged fleshy leaves emerging from the stem or surrounding the base of the stem, as seen in the iris (see figure below).

Link to Learning
Some aerial modifications of stems are tendrils and thorns. Tendrils are slender, twining strands that enable a plant (like a vine or pumpkin) to seek support by climbing on other surfaces. Thorns are modified branches appearing as sharp outgrowths that protect the plant; common examples include roses, Osage orange, and devil’s walking stick (see figure below).

Section Summary
The stem of a plant bears the leaves, flowers, and fruits. Stems are characterized by the presence of nodes (the points of attachment for leaves or branches) and internodes (regions between nodes).
Plant organs are made up of simple and complex tissues. The stem has three tissue systems: dermal, vascular, and ground tissue. Dermal tissue is the outer covering of the plant. It contains epidermal cells, stomata, guard cells, and trichomes. Vascular tissue is made up of xylem and phloem tissues and conducts water, minerals, and photosynthetic products. Ground tissue is responsible for photosynthesis and support and is composed of parenchyma, collenchyma, and sclerenchyma cells.
Primary growth occurs at the tips of roots and shoots, causing an increase in length. Woody plants may also exhibit secondary growth, or increase in thickness. In woody plants, especially trees, annual rings may form as growth slows at the end of each season. Some plant species have modified stems that help to store food, propagate new plants, or discourage predators. Rhizomes, corms, stolons, runners, tubers, bulbs, tendrils, and thorns are examples of modified stems.
Review Questions
Critical Thinking Questions
Glossary
- apical bud
- bud formed at the tip of the shoot
- axillary bud
- bud located in the axil: the stem area where the petiole connects to the stem
- bark
- tough, waterproof, outer epidermal layer of cork cells
- bulb
- modified underground stem that consists of a large bud surrounded by numerous leaf scales
- collenchyma cell
- elongated plant cell with unevenly thickened walls; provides structural support to the stem and leaves
- companion cell
- phloem cell that is connected to sieve-tube cells; has large amounts of ribosomes and mitochondria
- corm
- rounded, fleshy underground stem that contains stored food
- cortex
- ground tissue found between the vascular tissue and the epidermis in a stem or root
- epidermis
- single layer of cells found in plant dermal tissue; covers and protects underlying tissue
- guard cells
- paired cells on either side of a stoma that control stomatal opening and thereby regulate the movement of gases and water vapor
- internode
- region between nodes on the stem
- lenticel
- opening on the surface of mature woody stems that facilitates gas exchange
- node
- point along the stem at which leaves, flowers, or aerial roots originate
- parenchyma cell
- most common type of plant cell; found in the stem, root, leaf, and in fruit pulp; site of photosynthesis and starch storage
- periderm
- outermost covering of woody stems; consists of the cork cambium, cork cells, and the phelloderm
- pith
- ground tissue found towards the interior of the vascular tissue in a stem or root
- primary growth
- growth resulting in an increase in length of the stem and the root; caused by cell division in the shoot or root apical meristem
- rhizome
- modified underground stem that grows horizontally to the soil surface and has nodes and internodes
- runner
- stolon that runs above the ground and produces new clone plants at nodes
- sclerenchyma cell
- plant cell that has thick secondary walls and provides structural support; usually dead at maturity
- secondary growth
- growth resulting in an increase in thickness or girth; caused by the lateral meristem and cork cambium
- sieve-tube cell
- phloem cell arranged end to end to form a sieve tube that transports organic substances such as sugars and amino acids
- stolon
- modified stem that runs parallel to the ground and can give rise to new plants at the nodes
- tendril
- modified stem consisting of slender, twining strands used for support or climbing
- thorn
- modified stem branch appearing as a sharp outgrowth that protects the plant
- tracheid
- xylem cell with thick secondary walls that help transport water
- trichome
- hair-like structure on the epidermal surface
- tuber
- modified underground stem adapted for starch storage; has many adventitious buds
- vessel element
- xylem cell that is shorter than a tracheid and has thinner walls
