8.6 Superphylum Ecdysozoa: Nematodes and Tardigrades
Mary Ann Clark; Jung Choi; and Matthew Douglas
Learning Objectives
By the end of this section, you will be able to do the following:
- Describe the structural organization of nematodes
- Describe the importance of Caenorhabditis elegans in research
- Describe the features of Tardigrades
Superphylum Ecdysozoa
The superphylum Ecdysozoa contains an incredibly large number of species. This is because it contains two of the most diverse animal groups: phylum Nematoda (the roundworms) and phylum Arthropoda (the arthropods). The most prominent distinguishing feature of ecdysozoans is the cuticle—a tough, but flexible exoskeleton that protects these animals from water loss, predators, and other dangers of the external environment. One small phylum within the Ecdysozoa, with exceptional resistance to desiccation and other environmental hazards, is the Tardigrada. The nematodes, tardigrades, and arthropods all belong to the superphylum Ecdysozoa, which is believed to be monophyletic—a clade consisting of all evolutionary descendants from one common ancestor. All members of this superphylum periodically go through a molting process that culminates in ecdysis—the actual shedding of the old exoskeleton. (The term “ecdysis” translates roughly as “take off” or “strip.”) During the molting process, old cuticle is replaced by a new cuticle, which is secreted beneath it, and which will last until the next growth period.
Phylum Nematoda
The Nematoda, like other members of the superphylum Ecdysozoa, are triploblastic and possess an embryonic mesoderm that is sandwiched between the ectoderm and endoderm. They are also bilaterally symmetrical, meaning that a longitudinal section will divide them into right and left sides that are superficially symmetrical. In contrast with flatworms, nematodes are pseudocoelomates and show a tubular morphology and circular cross-section. Nematodes include both free-living and parasitic forms.
In 1914, N.A. Cobb said, “In short, if all the matter in the universe except the nematodes were swept away, our world would still be dimly recognizable, and if, as disembodied spirits, we could then investigate it, we should find its mountains, hills, vales, rivers, lakes and oceans represented by a thin film of nematodes…” To paraphrase Cobb, nematodes are so abundant that if all the non-nematode matter of the biosphere were removed, there would still remain a shadow of the former world outlined by nematodes!1 The phylum Nematoda includes more than 28,000 species with an estimated 16,000 being parasitic in nature. However, nematologists believe there may be over one million unclassified species.
The name Nematoda is derived from the Greek word “Nemos,” which means “thread,” and includes all true roundworms. Nematodes are present in all habitats, typically with each species occurring in great abundance. The free-living nematode, Caenorhabditis elegans, has been extensively used as a model system for many different avenues of biological inquiry in laboratories all over the world.
Morphology
The cylindrical body form of the nematodes is seen in the figure below. These animals have a complete digestive system with a distinct mouth and anus, whereas only one opening is present in the digestive tract of flatworms. The mouth opens into a muscular pharynx and intestine, which leads to a rectum and anal opening at the posterior end. The epidermis can be either a single layer of cells or a syncytium—a multinucleated tissue that in this case is formed by the fusion of many single cells. The cuticle of nematodes is rich in collagen and a polymer called chitin, which forms a protective armor outside the epidermis. The cuticle extends into both ends of the digestive tract, the pharynx, and the rectum. In the head, an anterior mouth opening is composed of three (or six) “lips” as well as teeth derived from the cuticle (in some species). Some nematodes may present other modifications of the cuticle such as rings, head shields, or warts. These external rings, however, do not reflect true internal body segmentation, which as we have seen is a hallmark of phylum Annelida. The attachment of the muscles of nematodes differs from that of most animals: they have a longitudinal layer only, and their direct attachment to the dorsal and ventral nerve cords creates a strong muscular contraction that results in a whiplike, almost spastic, body movement.
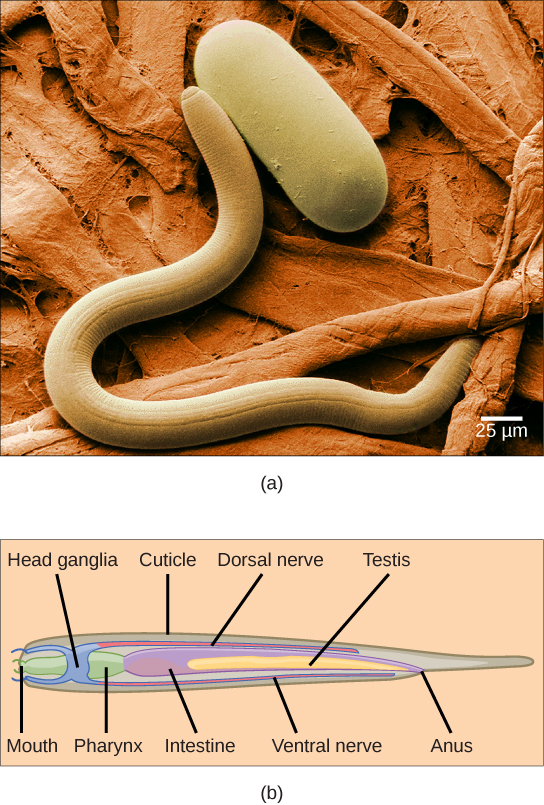
Excretory System
In nematodes, specialized excretory systems are not well developed. Nitrogenous wastes, largely in the form of ammonia, are released directly across the body wall. In some nematodes, osmoregulation and salt balance are performed by simple excretory cells or glands that may be connected to paired canals that release wastes through an anterior pore. In marine nematodes, the excretory cells are called renette cells, which are unique to nematodes.
Nervous system
Most nematodes have four longitudinal nerve cords that run along the length of the body in dorsal, ventral, and lateral positions. The ventral nerve cord is better developed than the dorsal and lateral cords. Nonetheless, all nerve cords fuse at the anterior end, to form a pharyngeal nerve ring around the pharynx, which acts as the head ganglion or the “brain” of the roundworm. A similar fusion forms a posterior ganglion at the tail. In C. elegans, the nervous system accounts for nearly one-third of the total number of cells in the animal!
Reproduction
Nematodes employ a variety of reproductive strategies ranging from monoecious to dioecious to parthenogenetic, depending upon the species. C. elegans is a mostly monoecious species with both self-fertilizing hermaphrodites and some males. In the hermaphrodites, ova and sperm develop at different times in the same gonad. Ova are contained in a uterus and amoeboid sperm are contained in a spermatheca (“sperm receptacle”). The uterus has an external opening known as the vulva. The female genital pore is near the middle of the body, whereas the male genital pore is nearer to the tip. In anatomical males, specialized structures called copulatory spicules at the tail of the male keep him in place and open the vulva of the female into which the amoeboid sperm travel into the spermatheca.
Fertilization is internal, and embryonic development starts very soon after fertilization. The embryo is released from the vulva during the gastrulation stage. The embryonic development stage lasts for 14 hours; development then continues through four successive larval stages with molting and ecdysis taking place between each stage—L1, L2, L3, and L4—ultimately leading to the development of a young adult worm. Adverse environmental conditions such as overcrowding or lack of food can result in the formation of an intermediate larval stage known as the dauer larva. An unusual feature of some nematodes is eutely: the body of a given species contains a specific number of cells as the consequence of a rigid developmental pathway.
Everyday Connection: C. elegans: The Model System for Linking Developmental Studies with Genetics
If biologists wanted to research how nicotine dependence develops in the body, how lipids are regulated, or observe the attractant or repellant properties of certain odors, they would clearly need to design three very different experiments. However, they might only need one subject of study: Caenorhabditis elegans. The nematode C. elegans was brought into the focus of mainstream biological research by Dr. Sydney Brenner. Since 1963, Dr. Brenner and scientists worldwide have used this animal as a model system to study many different physiological and developmental mechanisms.
C. elegans is a free-living nematode found in soil (see figure below). Only about a millimeter long, it can be cultured on agar plates (10,000 worms/plate!), feeding on the common intestinal bacterium Escherichia coli (another long-term resident of biological laboratories worldwide), and therefore can be readily grown and maintained in a laboratory. The biggest asset of this nematode is its transparency, which helps researchers observe and monitor changes within the animal with ease. It is also a simple organism with about 1,000 cells and a genome of only 20,000 genes. Its chromosomes are organized into five pairs of autosomes plus a pair of sex chromosomes, making it an ideal candidate with which to study genetics. Since every cell can be visualized and identified, this organism is useful for studying cellular phenomena like cell-to-cell interactions, cell-fate determinations, cell division, apoptosis (cell death), and intracellular transport.
Another tremendous asset is the short life cycle of this worm. It takes only three days to achieve the “egg to adult to daughter egg”; therefore, the developmental consequences of genetic changes can be quickly identified. The total life span of C. elegans is two to three weeks; hence, age-related phenomena are also easy to observe. There are two sexes in this species: hermaphrodites (XX) and males (XO). However, anatomical males are relatively infrequently obtained from matings between hermaphrodites, since their XO chromosome composition requires meiotic nondisjunction when both parents are XX. Another feature that makes C. elegans an excellent experimental model is that the position and number of the 959 cells present in adult hermaphrodites of this organism are constant. This feature is extremely significant when studying cell differentiation, cell-to-cell communication, and apoptosis. Lastly, C. elegans is also amenable to genetic manipulations using molecular methods, rounding off its usefulness as a model system.
Biologists worldwide have created information banks and groups dedicated to research using C. elegans. Their findings have led, for example, to better understanding of cell communication during development, neuronal signaling, and insight into lipid regulation (which is important in addressing health issues like the development of obesity and diabetes). In recent years, studies have enlightened the medical community with a better understanding of polycystic kidney disease. This simple organism has led biologists to complex and significant findings, growing the field of science in ways that touch the everyday world.
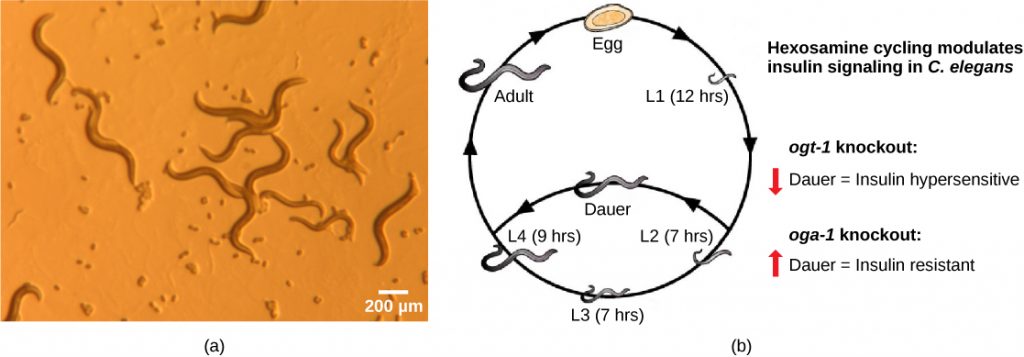
Parasitic Nematodes
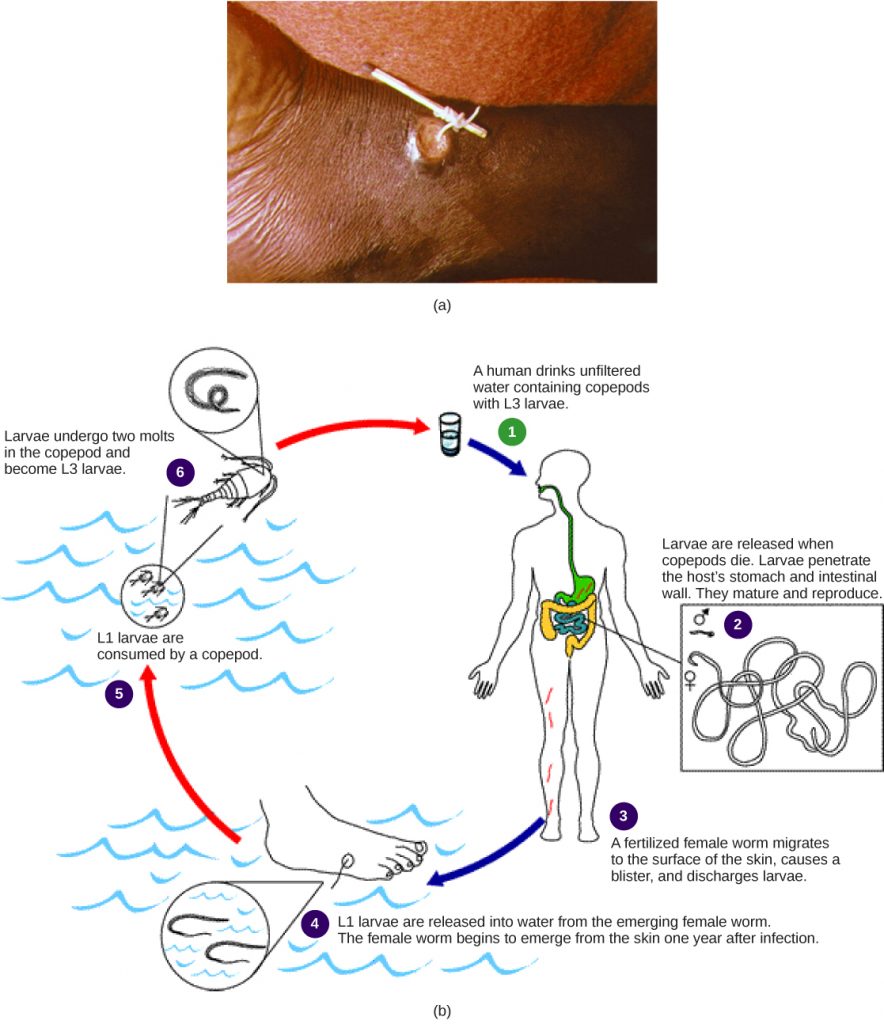
A number of common parasitic nematodes serve as prime examples of parasitism (endoparasitism). These economically and medically important animals exhibit complex life cycles that often involve multiple hosts, and they can have significant medical and veterinary impacts. Here is a partial list of nasty nematodes: Humans may become infected by Dracunculus medinensis, known as guinea worms, when they drink unfiltered water containing copepods, an intermediate crustacean host (see figure below). Hookworms, such as Ancylostoma and Necator, infest the intestines and feed on the blood of mammals, especially dogs, cats, and humans. Trichina worms (Trichinella) are the causal organism of trichinosis in humans, often resulting from the consumption of undercooked pork; Trichinella can infect other mammalian hosts as well. Ascaris, a large intestinal roundworm, steals nutrition from its human host and may create a physical blockage of the intestines. The filarial worms, such as Dirofilaria and Wuchereria, are commonly vectored by mosquitoes, which pass the infective agents among mammals through their blood-sucking activity. One species, Wuchereria bancrofti, infects the lymph nodes of over 120 million people worldwide, usually producing a non-lethal but deforming condition called elephantiasis. In this disease, parts of the body often swell to gigantic proportions due to obstruction of lymphatic drainage, inflammation of lymphatic tissues, and resulting edema. Dirofilaria immitis, a blood-infective parasite, is the notorious dog heartworm species.
Phylum Tardigrada
The tardigrades (“slow-steppers”) comprise a phylum of inconspicuous little animals living in marine, freshwater, or damp terrestrial environments throughout the world (see figure below). They are commonly called “water bears” because of their plump bodies and the large claws on their stubby legs. There are over 1,000 species, most of which are less than 1 mm in length. A chitinous cuticle covers the body surface and may be divided into plates. Tardigrades are known for their ability to enter a state called cryptobiosis, which provides them with resistance to multiple environmental challenges, including desiccation, very low temperatures, vacuum, high pressure, and radiation. They can suspend their metabolic activity for years and survive the loss of up to 99% of their water content. Their remarkable resistance has recently been attributed to unique proteins that replace water in their cells and protect their internal cell structure and their DNA from damage.

Morphology and Physiology
Tardigrades have cylindrical bodies, with four pairs of legs terminating in a number of claws. The cuticle is periodically shed, including the cuticular covering of the claws. The first three pairs of legs are used for walking, and the posterior pair for clinging to the substrate. A circular mouth leads to a muscular pharynx and salivary glands. Tardigrades feed on plants, algae, or small animals. Plant cells are pierced with a chitinous stylet and the cellular contents are then sucked into the gut by the muscular pharynx. Bands of single muscle cells are attached to the various points of the epidermis and extend into the legs to provide ambulatory movement. The major body cavity is a hemocoel, but there are no specialized circulatory structures for moving the blood, nor are there specialized respiratory structures. Malpighian tubules in the hemocoel remove metabolic wastes and transport them to the gut. A dorsal brain is connected to a ventral nerve cord with segmental ganglia associated with the appendages. Sensory structures are greatly reduced, but there is a pair of simple eyespots on the head, and sensory cilia or bristles concentrated toward the head end of the animal.
Reproduction
Most tardigrades are dioecious, and males and females each have a single gonad. Mating usually occurs at the time of a molt and fertilization is external. Eggs may be deposited in a molted cuticle or attached to other objects. Development is direct, and the animal may molt a dozen times during its lifetime. In tardigrades, like nematodes, development produces a fixed number of cells, with the actual number of cells being dependent on the species. Further growth occurs by enlarging the cells, not by multiplying them.
Section Summary
The defining feature of the Ecdysozoa is a collagenous/chitinous cuticle that covers the body, and the necessity to molt the cuticle periodically during growth. Nematodes are roundworms, with a pseudocoel body cavity. They have a complete digestive system, a differentiated nervous system, and a rudimentary excretory system. The phylum includes free-living species like Caenorhabditis elegans as well as many species of endoparasitic organisms such as Ascaris spp. They include dioeceous as well as hermaphroditic species. Embryonic development proceeds via several larval stages, and most adults have a fixed number of cells.
The tardigrades, sometimes called “water bears,” are a widespread group of tiny animals with a segmented cuticle covering the epidermis and four pairs of clawed legs. Like the nematodes, they are pseudocoelomates and have a fixed number of cells as adults. Specialized proteins enable them to enter cryptobiosis, a kind of suspended animation during which they can resist a number of adverse environmental conditions.
Review Questions
Critical Thinking Questions
Footnotes
- 1 Stoll, N. R., “This wormy world. 1947,” Journal of Parasitology 85(3) (1999): 392-396.
Glossary
- cuticle (animal)
- the tough, external layer possessed by members of the invertebrate class Ecdysozoa that is periodically molted and replaced
- Nematoda
- phylum of worm-like animals that are triploblastic, pseudocoelomates that can be free-living or parasitic
