99 Prokaryotic Cell Division
Learning Objectives
By the end of this section, you will be able to do the following:
- Describe the process of binary fission in prokaryotes
- Explain how FtsZ and tubulin proteins are examples of homology
Prokaryotes (bacteria and archaea) can undergo reproduction in which the cell increases to about double its original size and then undergoes division to form two daughter cells. For unicellular organisms, cell division is the only method to produce new individuals. In both prokaryotic and eukaryotic cells, the outcome of cell reproduction is a pair of daughter cells that are genetically identical to the parent cell. In unicellular organisms, daughter cells are individuals.
To achieve the outcome of cloned offspring, certain steps are essential. The genomic DNA must be replicated and then allocated into the daughter cells; the cytoplasmic contents must also be divided to give both new cells the cellular machinery to sustain life. As we’ve seen with bacterial cells, the genome consists of a single, circular DNA chromosome; therefore, the process of cell division is simplified. Karyokinesis is unnecessary because there is no true nucleus and thus no need to direct one copy of the multiple chromosomes into each daughter cell. This type of cell division is called binary (prokaryotic) fission.
Binary Fission
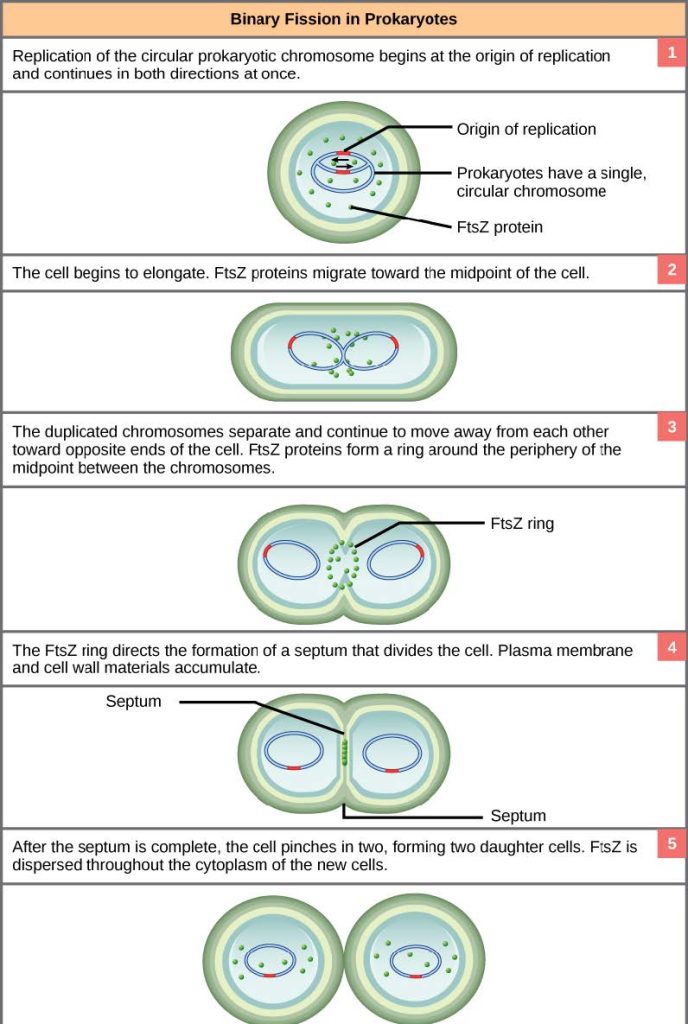
Due to the relative simplicity of the prokaryotes, the cell division process is a less complicated and much more rapid process than cell division in complex eukaryotes. Binary fission, meaning “division in half,” is the process by which prokaryotes reproduce from a parental cell into two daughter cells, each inheriting a complete genome and each a new individual. Binary fission also refers to the process of asexual reproduction of single-celled eukaryotes like amoeba, which involves mitosis, unlike prokaryotes like bacteria, which does not involve mitosis. As a review of the general information on cell division we discussed at the beginning of this chapter, recall that the single, circular DNA chromosome of bacteria occupies a specific location, the nucleoid region, within the cell (Figure 10.2). Although the DNA of the nucleoid is associated with proteins that aid in packaging the molecule into a compact size, there are no histone proteins and thus no nucleosomes in prokaryotes. The packing proteins of bacteria are, however, related to the cohesin and condensin proteins involved in the chromosome compaction of eukaryotes.
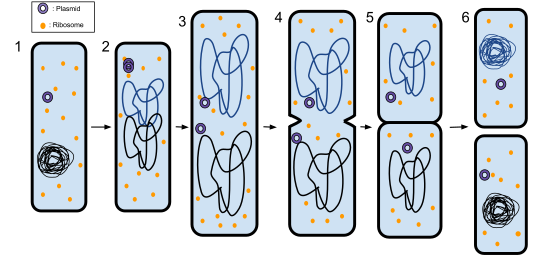
The bacterial chromosome is attached to the plasma membrane at about the midpoint of the cell. The starting point of replication, the origin, is close to the binding site of the chromosome to the plasma membrane (Figure 10.15a) and cells with plasmids proceed accordingly (Figure 10.15b). Replication of the DNA is bidirectional, moving away from the origin on both strands of the loop simultaneously. As the new double strands are formed, each origin point moves away from the cell wall attachment toward the opposite ends of the cell. As the cell elongates, the growing membrane aids in the transport of the chromosomes. After the chromosomes have cleared the midpoint of the elongated cell, cytoplasmic separation begins. The formation of a ring composed of repeating units of a protein called FtsZ (short for “filamenting temperature-sensitive mutant Z”) directs the partition between the nucleoids. Formation of the FtsZ ring triggers the accumulation of other proteins that work together to recruit new membrane and cell wall materials to the site. A septum is formed between the daughter nucleoids, extending gradually from the periphery toward the center of the cell. When the new cell walls are in place, the daughter cells separate.
Evolution Connection
Mitotic Spindle Apparatus
Did mitosis evolve from prokaryotic cell reproduction? There is a relatedness; however, we cannot observe evidence of extinct species’ cell divisions directly except to hypothesize. The precise timing and formation of the mitotic spindle is critical to the success of eukaryotic cell division. Prokaryotic cells, on the other hand, do not undergo karyokinesis and therefore have no need for a mitotic spindle. However, the FtsZ protein that plays such a vital role in prokaryotic cytokinesis is structurally and functionally very similar to tubulin, the building block of the microtubules, which make up the mitotic spindle fibers that are necessary for eukaryotic nuclear division. FtsZ proteins can form filaments, rings, and other three-dimensional structures that resemble the way tubulin forms microtubules, centrioles, and various cytoskeletal components. In addition, both FtsZ and tubulin employ the same energy source, GTP (guanosine triphosphate), to rapidly assemble and disassemble complex structures.
FtsZ and tubulin are considered to be homologous structures derived from common evolutionary origins. In this example, FtsZ is the ancestor protein to tubulin (an evolutionarily derived protein). While both proteins are found in extant organisms, tubulin function has evolved and diversified tremendously since evolving from its FtsZ prokaryotic origin. A survey of mitotic assembly components found in present-day unicellular eukaryotes reveals crucial intermediary steps to the complex membrane-enclosed genomes of multicellular eukaryotes (Table 10.3).
| Structure of genetic material | Division of nuclear material | Separation of daughter cells | |
|---|---|---|---|
| Prokaryotes | There is no nucleus. The single, circular chromosome exists in a region of cytoplasm called the nucleoid. | Occurs through binary fission. As the chromosome is replicated, the two copies move to opposite ends of the cell by an unknown mechanism. | FtsZ proteins assemble into a ring that pinches the cell in two. |
| Some protists | Linear chromosomes exist in the nucleus. | Chromosomes attach to the nuclear envelope, which remains intact. The mitotic spindle passes through the envelope and elongates the cell. No centrioles exist. | Microfilaments form a cleavage furrow that pinches the cell in two. |
| Other protists | Linear chromosomes wrapped around histones exist in the nucleus. | A mitotic spindle forms from the centrioles and passes through the nuclear membrane, which remains intact. Chromosomes attach to the mitotic spindle, which separates the chromosomes and elongates the cell. | Microfilaments form a cleavage furrow that pinches the cell in two. |
| Animal cells | Linear chromosomes exist in the nucleus. | A mitotic spindle forms from the centrosomes. The nuclear envelope dissolves. Chromosomes attach to the mitotic spindle, which separates the chromosomes and elongates the cell. | Microfilaments form a cleavage furrow that pinches the cell in two. |
